-
Importance de la structure bocagère sur les espèces animales présentes
 Hormis le caractéristiques faunistiques de la haie ou du couple haie-champ, d’autres paramètres doivent être pris en compte dans cette analyse : ce sont, en particulier, ceux qui concernent la structure bocagère, et qui peuvent être simplifiés en deux éléments :
Hormis le caractéristiques faunistiques de la haie ou du couple haie-champ, d’autres paramètres doivent être pris en compte dans cette analyse : ce sont, en particulier, ceux qui concernent la structure bocagère, et qui peuvent être simplifiés en deux éléments :
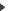 La surface des parcelles ou maillage :
La surface des parcelles ou maillage :
La densité de haies par hectare .
Le développement de l’écologie du paysage souligne le rôle de celui-ci dans la conservation ou le contrôle des pullulations d’espèces : ce n’est pas telle ou telle haie qui importe, ni dans certaines limites, leur longueur totale, c’est leur disposition dans l’espace, la continuité du réseau, les liens avec les zones incultes qu’il faut aussi considérer (Burel,1990, inter alia).
Ces deux éléments ont une influence sur les propriétés qualitative et quantitative de la faune. Une analyse comparative de l’alimentation des effraies sur la rive sud du Léman en 1926 –1927, puis en 1959 –1960, a montré une évolution du régime des oiseaux pendant cette période (Zelenka et Pricam, 1964 in Saint –Girons, 1976), les rongeurs des champs devenant de plus en plus nombreux (genre Microtus et Arvicola), et le pourcentage des insectivores diminuant. Or, dans le même temps, cette région, jadis bocagère, s’était transformée : les zones boisées diminuaient au bénéfice des cultures, et, surtout, la surface des parcelles augmentait, indiquant par la même la suppression de nombreux talus et haies vives. Cette exemple, ainsi que ceux de Saint-Girons (1965 et 1977), illustrent parfaitement l’ évolution de la faune du bocage vers une faune d’openfield .
-
La taille de la maille bocagère
Certains animaux installés dans les haies et les talus exploitent la parcelle plus ou moins profondément en fonction de leurs capacités de déplacement, laquelle est aussi liée à leur état physiologique. Aussi, les dimensions des parcelles peuvent-elles avoir des répercussions sur l’importance de la faune. Cette notion d’élargissement du bocage mérite d’être prise en compte : le dépassement d’une certaine surface de parcelles semble entraîner la disparitions d’animaux caractéristiques du bocage, les différentes microclimatiques et les effets de lisière étant moindre dans les grandes parcelles (Burel, 1990, inter alia).
La densité de haie
La relation entre maillage et avifaune est démontrée par le tableau suivant qui indique que la densité de couples reproducteurs varie en fonction de l’élargissement du bocage.
Bien que réalisé dans une autre région que celle concernée par ce rapport, les travaux de Grafeuille et al. (1981-1982) montrent que, en moyenne, dans un bocage à maille serrée (1ha), la densité, exprimée en couples d’espèces constantes, est 2,5 supérieure à celle observée dans un bocage à maille large (6 ha).
Les travaux de le Duc (1976), même si, comme le signale l’auteur, cette conclusion est à utiliser avec prudence, mentionnent que la maille bocagère peut-être élargie, sans influence sur les populations de chouettes hulottes (Stix aluco), à condition de ne pas dépasser une taille moyenne maximale de parcelles (5,8 ha pour la région considérée). Un maillage plus grand entraîne une brusque diminution de la densité de ce rapace.
Pour les reptiles, la population totale étant surtout fonction de la longueur de talus utilisable, elle diminue ave l’accroissement de dimensions du maillage (Saint-Girons et Duguy-1976).
Diversité et richesse spécifiques
Comme le montre le tableau précédent, l’élargissement de la maille bocagère ne provoque pas nécessairement de diminution du nombre total d’espèces de passereaux : des nicheurs de formations rases ou buissonnantes cohabitent.
En revanche, la situation est toute autre pour le nombre des espèces constantes et accessoires : 29 en bocage typique pour 20 en bocage à mailles élargies, et seulement 13 en zone ouverte (Eybert et Mahéo, 1975).
Pourtant, l’ouverture du paysage, si elle reste limitée, ne peut pas en faire diminuer, mais au contraire accroître la diversité écologique du milieu en faisant voisiner des zones de différentes tailles de grain ; si les suppressions de haies se poursuivent, la diversité de taille de grain reste constante pour un temps, puis décroît fortement lorsqu’il n’en subsiste plus que quelques unes dans un paysage devenu très ouvert (Braudy, 1984, in Courtiade, 1991 - Clavreul, 1984 - Courtiade, 1991).
Toutefois, dans les zones faiblement dégradées, une faune pauvre en espèces et peu abondante ne signifie pas qu’elle est sans intérêt : des nicheurs particuliers peuvent s’y trouver (Notteghem, 1986).
-
Les conséquences de la disparition des haies
Apparition d’espèces
Le remembrement en augmentant la taille des parcelles et en entraînant, par ces travaux connexes, une disparition des haies peut permettre à certaines espèces de s’y installer. Pour l’avifaune, notons les exemples de l’Alouette des champs et de la Linotte mélodieuse (Chandor et La Chapt, sans date - Collette, 1978 - Eybert et Mahéo, 1975).
Diminution ou disparition d’espèces
L’arasement des haies et des talus boisés entraîne, de facto, la disparition des espèces qui, à un moment ou l’autre de leur cycle de développement, sont liés à cette zone d’inculture. N’oublions pas, malgré tout, que toute diminution d’effectif n’est pas, nécessairement, à imputer à la seule action sur le milieu.
La disparition des reptiles(Saint-Girons et Duguy, 1976) ou des oiseaux (Clavreul, 1984 - Collette, 1978 et 1983) est protortionnelle à la longueur des haies arasées.
Pour l’avifaune, le nombre d’espèces est maximal dans une zone non arasée, alors qu’il est minimal dans la zone sans aucun talus 5eybert et Mahéo, 18975). Green et al. (1994) notent une réduction importante du nombre d’espèces à la suite d’un e coupe sévère des haies. Lang (1979) observe 22 espèces constantes ou accessoires dans le milieu fermé, et 20 dans le milieu ouvert. Les résultats de Biber et Biber (1990) ou de Clavreul (1984) indiquent que les parcelles situées dans le secteur bocager abritent un plus grand nombre d’espèces que les parcelles du secteur pauvre en haies.
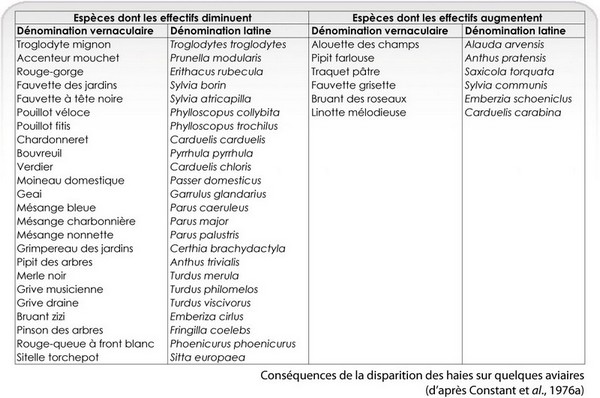
Lors de la diminution du nombre des talus, des espèces de passereaux, pour ne citer qu’elles, disparaissent totalement ou diminuent significativement. Corrélativement, d’autres augmentent leurs effectifs. Le détail des résultats est donné dans ce tableu.
Chez les Chiroptères, les travaux de Harouet (1991) semblent montrer un nombre d’individus plus important et une plus grande diversité spécifique sur le site noin remembré que sur le site remembré.
Pour le Putois, plus particulièrement lié aux bocages humides, les travaux connexes du remembrement (recalibrage des cours d’eau, arasement des haies et assèchements des zones marécageuses) entraînent une réduction notable des populations (Lodé, 1988). De plus, bien que n’étant pas seuls en cause, certains de ces facteurs accélèrent le disparition de la Loutre (Lodé, 1989).
Pullulation d’espèces
Le cloisonnement limiterait les territoires disponibles, et expliquerait, en partie, l’absence de pullulations de rongeurs dans les bocages (Saint-Girons, 1976). Le même auteur (1963 et 1965) signale que les pullulations cycliques des campagnols du genre Microtus y sont limités, et ne prennent jamais l’aspect catastrophique observé périodiquement dans le nord et l’est de la France.
-
Le cas particulier du remembrement
Sans oublier que les variations de populations ne sont quasi-jamais mono-factorielles, et que l’état initial du milieu semble prépondérant dans les conséquences de l’aménagement, l’action du remembrement se fait à plusieurs niveaux. Prenons l’exemple de l’avifaune :
Modification de la composition du peuplement : le remembrement laisse un milieu appauvri, et les pertes ne sont pas compensées, quantitativement et qualitativement, par l’installation de nouvelles espèces (Clavreul, 1984 - Collette, 1975, 1978 et 1983 - Eybert et Mahéo, 1975 - Lang, 1979),
Modification de la répartition spatiale : le nouveau peuplement n’occupe pas l’espace de la même façon que l’ancien, une majorité d’espèces se situant dans un espace restreint. Cette compression du peuplement met en évidence le rôle de refuge, de réservoir que peuvent jouer des micro-milieux au sein du bocage en période peu favorable (remembrement, hiver rude, ...),
modification du comportement : les passereaux semblent réagir au remembrement comme à un hiver exceptionnellement froid, la faible densité de la population s’accompagnant d’une réduction de l’activité vocale des mâles cantonnés (Collette, 1983).
Toutefois, lors d’un remembrement malgré la simplification du biotope, certaines espèces ont des populations tables. Cette situation peut cacher certaines réalités :
les faibles effectifs de l’espèce permettent à certains de ses individus d’occuper de nouvelles niches non encore exploitées.
l’espèce, même bien représentée, est numériquement stable, grâce à la grande adaptabilité de ses individus.
Cependant, la poursuite, avec excès, des travaux rompt l’équilibre : les espèces du premier groupe ne peuvent occuper de nouveaux sites, et c elles du second ne peuvent s’adapter indéfiniment aux modifications du milieux.
Comme l’écrit Collette (1983), à propos des passereaux bocagers, " le remembrement (...) apparaît donc actuellement une expérience nettement négative (...) ".
































