
Pôle Bocage
et Faune Sauvage


























-
Facteurs influençant la composition faunistique
 A côté des conditions climatiques générales, d’autres facteurs importants permettent d’expliquer la plus ou moins grande abondance des espèces dans une haie :
A côté des conditions climatiques générales, d’autres facteurs importants permettent d’expliquer la plus ou moins grande abondance des espèces dans une haie :
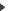 La structure de la végétation : des strates de végétations multiples, du fond du fossé à la cime des grands arbres.
La composition floristique : une flore diverse, selon les lieux et les saisons, permettant tous les régimes alimentaires herbivores, puis toutes les chaînes trophiques qui s’en suivent :
L’entretien des haies et des talus ;
Les connexions entre haies ;
La nature des cultures avoisinantes, des productions et techniques culturales variées, offertes par le système polyculture-élevage (Lebeau, 1979) ;
Les facteurs physiques de la « haie » : des microclimats divers (soleil, ombre, vent ou abri).
La structure de la végétation : des strates de végétations multiples, du fond du fossé à la cime des grands arbres.
La composition floristique : une flore diverse, selon les lieux et les saisons, permettant tous les régimes alimentaires herbivores, puis toutes les chaînes trophiques qui s’en suivent :
L’entretien des haies et des talus ;
Les connexions entre haies ;
La nature des cultures avoisinantes, des productions et techniques culturales variées, offertes par le système polyculture-élevage (Lebeau, 1979) ;
Les facteurs physiques de la « haie » : des microclimats divers (soleil, ombre, vent ou abri). -
Les caractéristiques de la haie qui influent sur la faune sauvage
Les haies boisées sont très complexes en elles-mêmes, différentes localement et d’une région à l’autre, et variables dans le temps de façon cyclique en fonction de leur entretien.
Leur rôle semble se manifester d’une double façon : par l’intermédiaire de la densité et de la hauteur du boisement, et comme facteur hétérogénéité favorable à la richesse du peuplement (Clavreul, 1984 –Constant, 1976 –Constant et al., 1976a –Saint-Girons, 1984 -Saint-Girons 1984 -Saint-Girons et al., 1986).La structure de la végétation
Du fait de leur largeur limitée, toutes les strates possibles de végétation peuvent être simultanément présentes dans les formations végétales des haies de cloisonnement.
Le peuplement avien (Constant et al. 1976a) ou micromammalien (Saint-Girons et al., 1986) semble réagir fortement à la structure de la végétation : des espèces sont infédoées, plus ou moins nettement, à une structure végétale ou à des strates de végétation, les vieilles souches et leurs excavations moussues représentant un site très important pour certaines d’entre elles. Comme l’écrivent Blondel et al. (1973) : « la diversité des peuplements d’oiseaux (…) est (…) directement fonction du degré de complexité structurale de la végétation, celle-ci pouvant alors être considérée comme la variable écologique fondamentale, les autres, intervenant indirectement par son intermédiaire ». Notteghem (1986) montre que le volume de végétation ligneuse est l’un des paramètres déterminants dans l’abondance totale de l’avifaune, et Coquillart (1987) que la hauteur de la végétation structure les peuplements avifaunistiques.
Les haies composés de trois strates (une arborescente dense, une arbustive, et une buissonnante dense) abriteront le plus grand nombre d’espèces, tant chez les reptiles (Saint-Girons et Duguy, 1976), les micromammifères (Saint-Girons et al.1976a –Osborne, 1984 –Parish et al., 1994 –Shalaway, 1985). Des résultats concernant l’avifaune sont consignés dans ce tableau.
 Pour les passereaux, l’important étant que la strate buissonnante basse soit bien garnie et fermée jusqu’au sol (Biber et Biber, 1980 –Notteghem, 1987), ils sont, dans leur majorité, plus sensibles à la disparition de la strate basse que haute. Lack (1988b) signale l’intérêt prédominant de la strate arbustive sur la strate arborée, pour la nidification des 11 espèces étudiées – notamment, le merle noir, l’accenteur mouchet, les grives draine et musicienne, et la fauvette grisette -, même si les deux sont nécessaires. Toutefois, pour les haies larges, si la situation est semblable aux formations forestières, le recouvrement arboré seul influerait négativement sur l’avifaune : en provoquant un excès de fermeture, il s’accompagne d’une diminution du nombre de niches, donc de la variété en oiseaux (Blondel et al., 1973 –Lebreton et al. 1987).Chez les micromammifères aussi, la simplification progressive de la haie se traduit par une diminution des effectifs et de la diversité du peuplement, mais la strate arboré ne semble jouer aucun rôle (Saint-Girons et al., 1986).Le mode et la périodicité d’entretien des haies et des talus modifient leur composition faunistique, ces pratiques provoquant, le plus souvent, une diminution de la diversité.Constant et al. (1976a) ont montré que les haies les plus âgées et les moins bien entretenues étaient les plus riches, l’émondage provoquant la disparition d’un certain nombre d’espèces, et une diminution des densités d’oiseaux. Parmi les espèces disparues, peuvent être notées la sitelle torchepot (Sitta eropaea), le pouillot véloce (Phulloscpus collybita), et la corneille (Corvus corone). Quant aux diminutions, citons les mésanges charbonnières (Parus major) et bleue (Parus caerulus), à la suite, semble-t-il, la disparition des fissures sur les branches les plus épaisses. A l’inverse, l’effectif d’étourneaux (Sturnus vulgaris) augmente nettement, alors que leur population n’est pas sans poser de problèmes dans le grand-ouest (Grarnet et Dubaille, 1983 –Douville de Franssu et al., 1991 –Pichot, 1982).Cependant, cette baisse de la diversité n’est pas générale à tous les animaux : chez les invertébrés arthropodes, une augmentation de certaines espèces est notée (INRA, 1976).
Pour les passereaux, l’important étant que la strate buissonnante basse soit bien garnie et fermée jusqu’au sol (Biber et Biber, 1980 –Notteghem, 1987), ils sont, dans leur majorité, plus sensibles à la disparition de la strate basse que haute. Lack (1988b) signale l’intérêt prédominant de la strate arbustive sur la strate arborée, pour la nidification des 11 espèces étudiées – notamment, le merle noir, l’accenteur mouchet, les grives draine et musicienne, et la fauvette grisette -, même si les deux sont nécessaires. Toutefois, pour les haies larges, si la situation est semblable aux formations forestières, le recouvrement arboré seul influerait négativement sur l’avifaune : en provoquant un excès de fermeture, il s’accompagne d’une diminution du nombre de niches, donc de la variété en oiseaux (Blondel et al., 1973 –Lebreton et al. 1987).Chez les micromammifères aussi, la simplification progressive de la haie se traduit par une diminution des effectifs et de la diversité du peuplement, mais la strate arboré ne semble jouer aucun rôle (Saint-Girons et al., 1986).Le mode et la périodicité d’entretien des haies et des talus modifient leur composition faunistique, ces pratiques provoquant, le plus souvent, une diminution de la diversité.Constant et al. (1976a) ont montré que les haies les plus âgées et les moins bien entretenues étaient les plus riches, l’émondage provoquant la disparition d’un certain nombre d’espèces, et une diminution des densités d’oiseaux. Parmi les espèces disparues, peuvent être notées la sitelle torchepot (Sitta eropaea), le pouillot véloce (Phulloscpus collybita), et la corneille (Corvus corone). Quant aux diminutions, citons les mésanges charbonnières (Parus major) et bleue (Parus caerulus), à la suite, semble-t-il, la disparition des fissures sur les branches les plus épaisses. A l’inverse, l’effectif d’étourneaux (Sturnus vulgaris) augmente nettement, alors que leur population n’est pas sans poser de problèmes dans le grand-ouest (Grarnet et Dubaille, 1983 –Douville de Franssu et al., 1991 –Pichot, 1982).Cependant, cette baisse de la diversité n’est pas générale à tous les animaux : chez les invertébrés arthropodes, une augmentation de certaines espèces est notée (INRA, 1976).Composition floristique
D’après certains auteurs (Blondel et al. 1973 – Lebreton et al.1987), la composition floristique n’est pas, pour les oiseaux nicheurs, un facteur déterminant de la constitution du biotope avien.Il apparaît cependant que l’influence de ce facteur a été assez peu étudié et est difficile à dissocier de l’influence de la structure de la végétation. Constant et al. (1976a) ont étudié l’importance de la composition floristique (tableau).

Il semblerait que la richesse qualitative et quantitative de l’avifaune soient liées à la diversité végétale.
Ces résultats sont toutefois à considérer avec précaution car il apparaît que la structure végétale des trois types de talus étudiés ne soit pas identique. Il est donc possible que le résultat obtenu soit dû à une combinaison des facteurs composition floristique et structure végétale.Connexions
Pour l’avifaune, Constant et al. (1976a) et Notteghem (1986), par exemple, signalent que les zones privilégiées de cantonnement sont les intersections de talus, un gradient décroissant étant observé depuis les connexions des haies : les premiers y dénombrent 68,7 % des oiseaux. Cet « effet-carrefour » est particulièrement marqué chez certaines espèces : accenteur mouchet (Prunella modularis), pouillot véloce, mésange bleue et pinson des arbres (Fringilla coelebs).Ce phénomène s’expliquerait par (Constant et al. 1976a –Lack, 1988a) :
La compacité du territoire à défendre : près d’une intersection, sur une plus petite surface, la longueur des haies est supérieure à celle obtenue avec une surface identique dans une zone linéaire. Plus compacte, la zone est plus aisée à défendre ;
L’accroissement de la nourriture qui se trouve dans les zones proches des haies ;
Une augmentation des sites de protection contre les intempéries venant de différentes directions :
Une augmentation des zones de refuge en cas de dérangement ou d’attaques de prédateurs :
Une plus grande richesse de la haie à cause de la culture moins aisée dans le coin du champ. -
Les structures adjacentes influent sur la faune sauvage
 De nombreux travaux font état des variations de la composition faunistique en fonction des différents types d’utilisation des champs. Tel est le cas des micromammifères (Constant et al. 1976b) ou des oiseau (Arnold 1983 –Clavreul, 1984 –Constant et al. 1976a –Green et al. 1994 –Lakhani, 1994 –Parish et al. 1994). Ces derniers auteurs signalent que la pâture permanente, plutôt que les terres arables dans les champs adjacents, augmente la richesse en espèces d’oiseaux, particulièrement en étant associée avec de larges haies.En outre, les remaniements occasionnés par les travaux du sol (labour, passage du rotovator) obligent certains groupes, tel que les micromammifères (Constant, 1976 –Constant et al. 1976b), à effectuer des cycles de déplacement saisonnier.
De nombreux travaux font état des variations de la composition faunistique en fonction des différents types d’utilisation des champs. Tel est le cas des micromammifères (Constant et al. 1976b) ou des oiseau (Arnold 1983 –Clavreul, 1984 –Constant et al. 1976a –Green et al. 1994 –Lakhani, 1994 –Parish et al. 1994). Ces derniers auteurs signalent que la pâture permanente, plutôt que les terres arables dans les champs adjacents, augmente la richesse en espèces d’oiseaux, particulièrement en étant associée avec de larges haies.En outre, les remaniements occasionnés par les travaux du sol (labour, passage du rotovator) obligent certains groupes, tel que les micromammifères (Constant, 1976 –Constant et al. 1976b), à effectuer des cycles de déplacement saisonnier. -
Les acteurs physiques qui influent sur la faune sauvage
Microclimat
Les bioclimatologiste (INRA, 1976) ont montré que des gradients de vitesse de vent et de températures existaient au sein de la parcelle, et pouvaient agir sur :
L’installation de différents animaux en des lieux choisis en fonction de leurs « préférences » pour certaines conditions de température, d’humidité, d’ensoleillement, etc. ; ces conditions de milieu peuvent intervenir directement sur les espèces concernées ou indirectement par l’abondance de leurs proies par exemple.
Le développement des populations en place avec une rapidité plus ou moins grande, en fonction des températures.Sol, eau
Ces facteurs peuvent aussi agir sur la répartition des animaux. Constant (1976) signale que le substrat ou le niveau piézométrique influence la répartition des espèces de rongeurs. -
Les variations périodiques annuelles influent sur la faune sauvage
L’hétérogénéité spatiale globale se complique par les variations intra- et interannuelles de la répartition des espèces. Les conditions dans lesquelles se déroulent chaque année la colonisation des milieux dépendent, en effet, de nombreux facteurs parmi lesquels :
les conditions météorologiques au cours de la période de reproduction,
les conditions d’hivernage, et
le déroulement de la migration.Pour l’avifaune, le nombre d’espèces utilisant les haies est influencé par les caractéristiques du milieu environnant, et ce plus fortement en été qu’en hiver (Arnold, 1983). Clavreul (1984) montre que certaines espèces peuvent ne pas nicher régulièrement en un point donné. Souvent négligeable dans les grands massifs forestiers tempérés, cette influence peut être plus marquée sur des zones de surface plus réduite.





